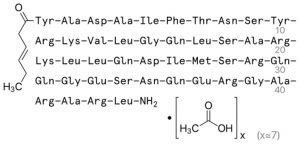
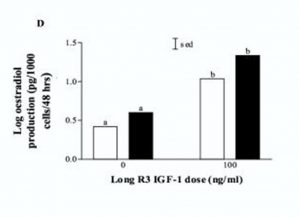
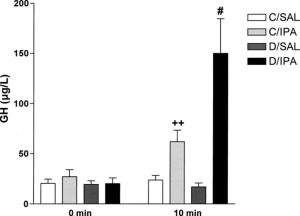
Bremelanotide is a 7 amino acid (AA) analogue of α melanocyte-stimulating hormone (α-MSH) used in scientific research studies as a treatment for hyposexual sexual desire disorder (HSDD) in women1. As a peptide, bremelanotide is injected subcutaneously, although intranasal administration has shown efficacy in stimulating sexual arousal in female vitro test subjects. Male vitro test subjects with sexual dysfunction also demonstrate increased sexual arousal and erectile responses after subcutaneous bremelanotide injection, including subjects for whom Viagra was ineffective3. In addition to its beneficial role in sexual dysfunction studies, bremelanotide has excellent utility in many different fields of neuroscience research.
DISCLAIMER: All products from Blue Sky Peptide are for laboratory research only and are not suitable for human consumption. Any other use violates the terms and conditions for purchase. Bremelanotide should only be used in carefully controlled laboratory conditions with appropriate personal protective equipment. If exposure occurs, immediately rinse the affected area and seek medical help if symptoms present.
Bremelanotide is a melanocortin receptor (MCR) agonist that preferentially interacts with the melanocortin receptor subtype 4 (MC4R)4. The melanocortins and their receptors were first identified as pituitary factors involved in skin darkening in frogs. Eventually, however, the exploration of the melanocortin system extended to mammalian systems5. The melanocortin system contributes to skin pigmentation, inflammation, appetite, memory, and sexual behavior5-8.
The different MCR subtypes play distinct roles and are differentially expressed across the body. Melanocortins, also known as melanocyte-stimulating hormones (MSH), are all small peptide hormones derived from proopiomelanocortin and serve as the primary agonists of MCRs along with adrenocorticotropic hormone (ACTH)9. MSH peptides come in 3 types: α-MSH, β-MSH, γ-MSH.
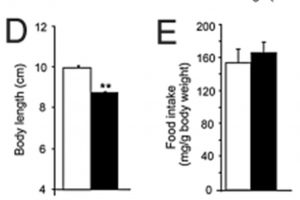
MC4R is primarily localized in the brain and is considered a neural MCR10. One of the primary roles of MC4R is regulating food intake by mediating the effects of leptin11. Administering α-MSH reduces feeding in obese rats, especially when applied directly to the paraventricular nucleus of the hypothalamus (PVN), a critical autonomic control center of the brain involved with stress responses, growth, and reproduction12-14. In addition to food intake, MC4R plays a role in other processes like glucose and lipid homeostasis, disease-associated lean body mass wasting, and cardiovascular function10.
Reproductive behavior is also intrinsically tied to MC4R and its ligands. Mice with reduced or absent MC4R function exhibit several sexual dysfunctions15-17. Administration of MC4R agonist reversed these effects in some studies and caused erections and ejaculations in some animal models after central nervous system administration10,18.
Considering the breadth of life-essential roles the melanocortin system has, an MC4R-specific agonist like bremelanocortin is valuable for many lines of neuroscience research. In addition, bremelanocortin is extensively characterized in the literature and is well-tolerated by many species, including rats and humans. Therefore, bremelanocortin is, and has been, a powerful research tool.
References:
- Mayer D, Lynch SE. Bremelanotide: New Drug Approved for Treating Hypoactive Sexual Desire Disorder. Ann Pharmacother. 2020;54(7):684-690.
- Clayton AH, Althof SE, Kingsberg S, et al. Bremelanotide for female sexual dysfunctions in premenopausal women: a randomized, placebo-controlled dose-finding trial. Womens Health (Lond). 2016;12(3):325-337.
- Rosen RC, Diamond LE, Earle DC, Shadiack AM, Molinoff PB. Evaluation of the safety, pharmacokinetics and pharmacodynamic effects of subcutaneously administered PT-141, a melanocortin receptor agonist, in healthy male subjects and in patients with an inadequate response to Viagra. Int J Impot Res. 2004;16(2):135-142.
- Pfaus JG, Sadiq A, Spana C, Clayton AH. The neurobiology of bremelanotide for the treatment of hypoactive sexual desire disorder in premenopausal women. CNS Spectr. 2021:1-9.
- Wikberg JE, Muceniece R, Mandrika I, et al. New aspects on the melanocortins and their receptors. Pharmacol Res. 2000;42(5):393-420.
- Shen Y, Fu WY, Cheng EY, Fu AK, Ip NY. Melanocortin-4 receptor regulates hippocampal synaptic plasticity through a protein kinase A-dependent mechanism. J Neurosci. 2013;33(2):464-472.
- Catania A, Lonati C, Sordi A, Carlin A, Leonardi P, Gatti S. The melanocortin system in control of inflammation. ScientificWorldJournal. 2010;10:1840-1853.
- Ellacott KL, Cone RD. The role of the central melanocortin system in the regulation of food intake and energy homeostasis: lessons from mouse models. Philos Trans R Soc Lond B Biol Sci. 2006;361(1471):1265-1274.
- Gantz I, Fong TM. The melanocortin system. Am J Physiol Endocrinol Metab. 2003;284(3):E468-474.
- Tao YX. The melanocortin-4 receptor: physiology, pharmacology, and pathophysiology. Endocr Rev. 2010;31(4):506-543.
- Kask A, Rago L, Wikberg JE, Schioth HB. Evidence for involvement of the melanocortin MC4 receptor in the effects of leptin on food intake and body weight. Eur J Pharmacol. 1998;360(1):15-19.
- Ferguson AV, Latchford KJ, Samson WK. The paraventricular nucleus of the hypothalamus - a potential target for integrative treatment of autonomic dysfunction. Expert Opin Ther Targets. 2008;12(6):717-727.
- Hansen MJ, Ball MJ, Morris MJ. Enhanced inhibitory feeding response to alpha-melanocyte stimulating hormone in the diet-induced obese rat. Brain Res. 2001;892(1):130-137.
- Mountjoy KG, Mortrud MT, Low MJ, Simerly RB, Cone RD. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol Endocrinol. 1994;8(10):1298-1308.
- Sandrock M, Schulz A, Merkwitz C, Schoneberg T, Spanel-Borowski K, Ricken A. Reduction in corpora lutea number in obese melanocortin-4-receptor-deficient mice. Reprod Biol Endocrinol. 2009;7:24.
- Irani BG, Xiang Z, Moore MC, Mandel RJ, Haskell-Luevano C. Voluntary exercise delays monogenetic obesity and overcomes reproductive dysfunction of the melanocortin-4 receptor knockout mouse. Biochem Biophys Res Commun. 2005;326(3):638-644.
- Van der Ploeg LH, Martin WJ, Howard AD, et al. A role for the melanocortin 4 receptor in sexual function. Proc Natl Acad Sci U S A. 2002;99(17):11381-11386.